Projects
-
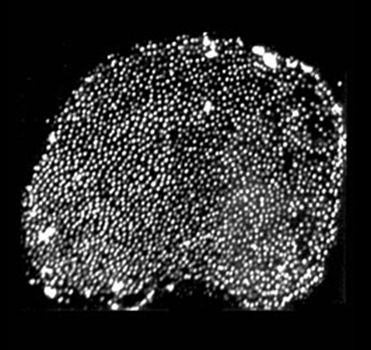
While a postdoc in Jim Hudspeth's lab, Peter Barr-Gillespie developed the “twist-off” method for isolating hair bundles, the sensory organelle of hair cells, from the frog inner ear (Gillespie & Hudspeth, 1991). The Barr-Gillespie lab has applied the purification method to bundles from inner ears of rat (Dumont et al., 2002), mouse (Holt et al., 2002), and chicken (Shin et al., 2007). Moreover, we can use biochemical methods like immunoblotting, immunoprecipitation, photoaffinity labeling, and mass spectrometry to characterize proteins from isolated bundles (Walker et al., 1993; Gillespie et al., 1993; Yamoah & Gillespie, 1996; Hasson et al., 1997; Yamoah et al., 1998; Dumont et al., 2001; Hill et al., 2006).
Because hair bundles account for perhaps 1% of the total protein in an auditory or vestibular epithelium, using the bundle preparation allows us to focus on protein variants, interactions, and mechanisms that are found in the mechanosensory organelle. We expect that the combination of the bundle preparation and mass spectrometry will allow us to identify key molecules involved in bundle function, including transduction.
-
Although we developed the "twist-off" method for isolating hair bundles many years ago (Gillespie & Hudspeth, 1991), protein sequencing technologies have only recently matured sufficiently to allow detection of the small amounts of protein in hair bundles. In particular, we now use shotgun proteomics methods with mass spectrometry detection and identification (Domon & Aebersold, 2006) to detect and measure hair bundle proteins (Shin et al., 2007).
Mass spectrometry allows us to detect proteins as scarce as 1/1000th the concentration of the most abundant protein, actin. By developing methods to purify components of the bundle, most significantly their plasma membranes, we should be able to detect proteins far less abundant, perhaps even the transduction channel itself. Mass spectrometry analysis opens up a vast new world of hair-bundle biochemistry, which the Barr-Gillespie lab is beginning to explore.
-
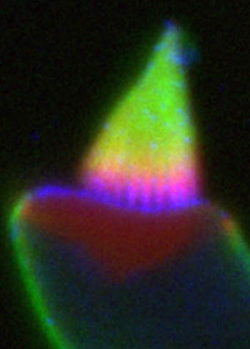
Little is known about the composition and physical properties of the plasma membrane of the hair bundle. Several lipids have remarkably asymmetric distributions in stereocilia, suggesting the presence of unique lipid domains that may have functional significance.
We have shown that the lipid phosphatidylinositol-4,5-bisphosphate (PIP2) is localized above the ankle links in stereocilia, being absent from the taper region and apical surface. A lipid phosphatase, Ptprq, shows the reciprocal distribution (Hirono et al., 2004), suggesting that the phosphatase may contribute to PIP2 localization. We are presently determining the location of enzymes involved in PIP2 biosynthesis in hair bundles.
Recent data show that a polysialyated form of GM1 ganglioside is restricted to the same area as Ptprq. Present experiments are focused on determining causality of localization of this lipid and the enzyme Ptprq.
Finally, we have adapted the hair bundle purification method to allow us to characterize the composition of the bundle plasma membrane using lipid mass spectrometry. We are interested whether the membrane composition is sufficiently distinct to alter the physical properties of the bundle membrane.
-
When examining the list of major bundle proteins identified by mass spectrometry, we were struck by the number that were involved in energy metabolism (Shin et al., 2007). The brain isoform of creatine kinase was the second most abundant protein, after actin; enzymes participating in glycolysis were also present at substantial levels.
These observations fit in with previous calculations that showed that ATP consumption in hair bundles could be very high. At a density in bundles of 2000 molecules per square micron, the plasma-membrane Ca2+-ATPase (PMCA) is capable of pumping out all Ca2+ that enters a stereocilium during mechanotransduction, which can be >1 mM/s (Yamoah et al., 1998). Because an ATP molecule is consumed for every calcium ion removed, bundles may consume a prodigious amount of ATP under some conditions.
No mitochondria are present in hair bundles, suggesting that bundles rely on ATP diffusion or other methods for maintaining ATP levels. We have shown that diffusion from cell bodies likely cannot maintain high ATP in bundles, suggesting that creatine kinase and glycolytic enzymes are targeted to bundles specifically to maintain ATP levels high enough for myosins and other bundle ATPases.
Download the Barr-Gillespie hair bundle ATP pathway diagram. (97 KB)
-

Unconventional myosins play critical roles in hair bundle function, not surprising given the prominence of the specialized actin cytoskeleton of the bundle. Mutations in five myosin genes, including myosins -1a, -3a, -6, -7a, and -15a, lead to deafness and balance dysfunction in humans, mice, or fish. Moreover, localization and chemical genetics experiments have implicated a sixth member of the family, myosin-1c, as being an essential part of the adaptation motor.
Our lab has adapted methods pioneered by Kevan Shokat's lab (Shah et al., 1997) to allow selective inhibition of a myosin-1c allele (Gillespie et al., 1999). We have introduced mutant alleles into the mouse genome using either transgenic (Holt et al., 2002) or gene targeting (Stauffer et al., 2005) approaches. Examination of mechanotransduction in these mice led us to propose not only that myosin-1c was the motor responsible for slow adaptation, but also that it participated in fast adaptation.
Although we are very interested in applying similar methods to other myosins important for hair cell function, technical hurdles have hindered development of myosin-6 and myosin-7a mutant alleles. Nevertheless, we suggest that our approach is general to the myosins and thus could be used with myosins for which we have only poor functional understanding.
-
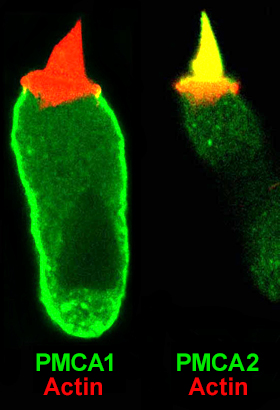
Ca2+ plays a central role in transduction, regulating slow (Eatock et al., 1987) and fast (Crawford et al., 1991) adaptation, as well as tip-link regeneration (Zhao et al., 1996). Although Ca2+ entering through transduction channels is rapidly bound by mobile Ca2+ buffers (Ricci et al., 1998), most is pumped out by the plasma-membrane Ca2+ ATPase (PMCA) (Lumpkin & Hudspeth, 1998). Moreover, although mobile Ca2+ buffers control spatial gradients of Ca2+ when transduction channels are open, PMCA activity sets the channels-closed level of Ca2+, which determines how much influence entering Ca2+ can have on adaptation. Finally, PMCA is likely to be the major ATP drain in bundles (Shin et al., 2007).
We (Dumont et al., 2001; Hill et al., 2006) and others (e.g., Grati et al., 2006) have shown that the Ca2+ pump of mammalian hair bundles is PMCA2w/a. Mice with null mutations in PMCA2 are deaf (Street et al., 1998; Kozel et al., 1998), illustrating PMCA2's importance.
Our future goal is to determine how PMCA2w/a is targeted to stereocilia membranes and how the cell determines how to control protein and activity levels of the pump, presumably providing precise control over the rate of Ca2+ pumping from stereocilia. Moreover, we intend to test the hypothesis that the calcium pump can significantly elevate the concentration of Ca2+ close to hair bundles.
-
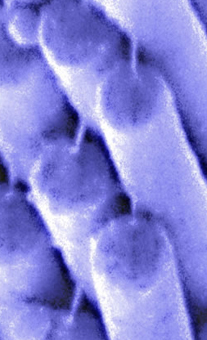
First identified by Jim Pickles in 1984, tip links are extracellular links connecting adjacent stereocilia along the mechanically sensitive axis of the hair bundle (Pickles et al., 1984). Tip links are essential for transduction (Assad et al., 1991) and are thought to be or are in series with the gating springs, the mechanical elements that directly gate transduction channels (Corey & Hudspeth, 1983).
In collaboration with Bechara Kachar, an outstanding microscopist at the NIH, we showed that the tip link is a helical dimer that appears relatively stiff (Kachar et al., 2000). In subsequent work in collaboration with Uli Mueller, an investigator at Johns Hopkins University, we suggested that the tip link was at least partially composed of a member of the cadherin superfamily known as cadherin 23 (Siemens et al., 2004). Cadherin 23 was already well known as a "deafness gene" (DiPalma et al, 2001; Bolz et al., 2001; Wilson et al., 2001; Bork et al., 2001), but its role in hair-cell function has not been easy to tease out. Other groups have indicated that protocadherin 15 is another component of the tip link (Ahmed et al., 2006).
The Barr-Gillespie lab has shown that tip links have the amazing ability to regenerate following chemical ablation (Zhao et al., 1996); mechanotransduction returns simultaneously, albeit with minimal adaptation. The ongoing molecular identification of the tip link will allow us to revisit these experiments and work out the mechanism by which a hair bundle strings its tip link, positioning it in the proper orientation and subjecting it to sufficient tension to maintain transduction channels at their optimally sensitive points.
-
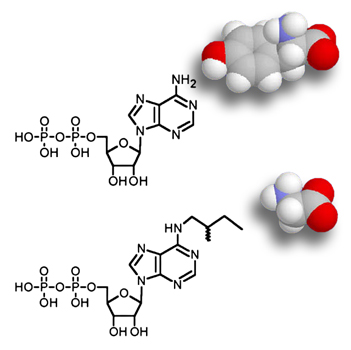
To better understand how a single protein participates in transduction, we have adapted a conceptual strategy that allows us to directly determine the role or location of a protein of interest. We alter a protein (for example, a myosin) in a way that has little or no effect on its activity (single amino-acid substitution or tagging), yet sensitizes it to an acutely-applied probe, such as an inhibitor or tag-binding reagent. We express the altered protein in hair cells, and then use the probe to acutely inhibit, activate, or detect the protein of interest. If the probe is both selective and specific for the altered/mutant protein — that is, there is no interaction observed in three negative controls: mutant hair cells without, wild-type cells without, and wild-type cells with the probe — then we can confidently conclude that the protein is an integral part of the mechanism of interest.
Applications include:
- specific inhibition or activation of sensitized mutant myosins using ADP and ATP analogs to identify roles in transduction or adaptation (chemical genetics)
- specific inhibition of tetracysteine-tagged myosins by light inactivation (FlAsH FALI)
- simultaneous delivery of two tagged proteins to assay protein-protein interactions by crosslinking, FRET, or coimmunoprecipitation
We originally applied this strategy to determining the roles of myosin-1c in hair cells. We are presently applying this approach to the hair-bundle Ca2+ pump and to candidates for the transduction channel itself. Moreover, we are planning similar experiments to establish definitively whether Cdh23 is indeed the tip link.
This method allows us to test the role of a protein in mechanotransduction in the protein's natural context — a live, functioning hair cell. We believe that this strategy allows us to better understand the roles of specific proteins in hair-cell transduction and adaptation.